goto
end
click here to download PDF verson
of this article
This article and/or assocatied PDF file may only be posted
for a limited time, due to web space.
The URL for this page is http://tellme1st.net/israels-ekklesia/Genes_in_Palestinians/Genes_in_Palestinians.html
HLA Genes in Palestinians
ABSTRACT: The genetic profile of Palestinians has, for the
first time, been studied by using human leukocyte antigen (HLA) gene variability
and haplotypes. The comparison with other Mediterranean populations by
using neighbor-joining dendrograms and correspondence analyses reveal that
Palestinians are genetically very close to Jews and other Middle East populations,
including Turks (Anatolians), Lebanese, Egyptians, Armenians and Iranians.
Archaeologic and genetic data support that both Jews and Palestinians came
from the ancient Canaanites, who extensively mixed with Egyptians, Mesopotamian
and Anatolian peoples in ancient times. Thus, Palestinian- Jewish rivalry
is based in cultural and religious, but not in genetic, differences. The
relatively close relatedness of both Jews and Palestinians to western Mediterranean
populations reflects the continuous circum-Mediterranean cultural and gene
flow that have occurred in prehistoric and historic times. This flow overtly
contradicts the demic diffusion model of western Mediterranean populations
substitution by agriculturalists coming from the Middle East in the Mesolithic-Neolithic
transition. Human Immunology 62, 889-900 (2001). ã American Society
for Histocompatibility and Immunogenetics, 2001. Published by Elsevier
Sciece Inc.
KEYWORDS: Palestinians; Macedonians; Greeks; Mediterraneans;
Berbers; Moroccans; Albanians; Turks; Iranians; Egyptians; Jews; Lebanese.
This work is dedicated to all Palestinians and Jews who
are suffering war. From de Department of Immunology and Molecular Biology
(A.A.-V., C.S, J.M., E.G.-C., L.A., P.V., J.M.-L.), H. 12 de Octubre, Universidad
Complutense, Madrid, Spain, and Laboratories & Blood Bank (N.E., A.R.),
El-Shifa Hospital, Gaza, Palestine. Address reprint request to Antonio
Arnaiz-Villena, Departamento de Inmunología y Biología Molecular,
H. 12 de Octubre, Universidad Complutense, Carretera Andalucía.
28041 Madrid, Spain; Tel: +34 (91) 390 8315; Fax: +34 (91) 390 8399; E-mail:
aarnaiz@eucmax.sim.ucm.es, Web address: http://chopo.pntic.mec.es/biolmol.
Received May 16, 2001; accepted June 15, 2001. Human Immunology 62, 889-900
(2001) ã American Society for Histocompatibility and Immunogenetics,
2001 Published by Elsevier Science Inc 0198-8859/01/$-see front matter
S0198-8859(01)00288-9
The Origin of Palestinians and Their Genetic Relatedness
With Other Mediterranean Populations
by Antonio Arnaiz-Villena, Nagah Elaiwa, Carlos
Silvera, Ahmed Rostom, Juan Moscoso, Eduardo Gómez-Casado, Luis
Allende, Pilar Varela, and Jorge Martínez-Laso
INTRODUCTION
The highly polymorphic human leukocyte antigen (HLA) system has been
validated as useful for distinguishing and/or relating populations (and
individuals) in many research studies since the first International HLA
Anthropology Workshop [1] and in all the subsequent International Workshops.
HLA gene frequencies correlate with geographically related populations.
The existence or absence of gene flow among neighbouring ethnic groups
may be assessed with the study of HLA frequencies and the corresponding
genetic distances and haplotypes [2, 3].
Most ancient recorded inhabitants of Palestine are named Canaanites
(3rd millennium BC or more ancient). They became urbanized and lived in
city-states, one of which was Jericho. Palestines location at the center
of routes linking three continents made it the meeting place for religious
and cultural influences from Egypt, Syria, Mesopotamia, and Anatolia. During
the second millennium BC, Egyptian hegemony and Canaanite autonomy were
constantly challenged by such ethnically diverse invaders as the Amorites,
Hittites, and Hurrians from Anatolia and the East. These invaders, however,
were defeated by the Egyptians and absorbed by the Canaanites, who at that
time may have numbered about 200,000. Egyptian power began to weaken, and
new invaders or autochthonous people appeared or made themselves noticeable
[4]. The Israelites, a confederation of Hebrew tribes, finally defeated
most of the Canaanites (1125 BC) but found the struggle with the Philistines
(Palestinians) more difficult. Philistines had established an independent
state on the southern coast of Palestine and also controlled the Canaanite
town of Jerusalem. The sea people contributed to the fall of the Anatolian
Hittites and other Middle East people by 1200 BC and apparently seriously
threatened Egypt [5]. The Philistines have been included among the sea
people invaders. However, it is doubtful that big amounts of people entered
nowadays Anatolia and Palestine; a new iron technology probably was taken
de novo by some autochthonous Canaanite tribes that acquired superiority.
In fact, the Palestinians are nowadays thought to come from the Egyptian
garrisons that were abandoned to their own fate on the Canaan land by 1200
years BC (Figure 1) and had to manage to construct or reinforce or rebuilt
some ancient Canaanite city-states, together with the old autochthonous
tribes [6]. Otherwise, the ancient Palestinians might have come from Crete
or its empire [7]. Israelites could also stem from autochthonous Canaanite
tribes that were agglutinated by a group of people led by Moses to fight
against other Canaanites, including Philistines and finally set up ancient
Israel [6- 8]. By 1000 BC, and after warring with Philistines and other
Canaanites, an Israelite state was founded by king Saul [6].
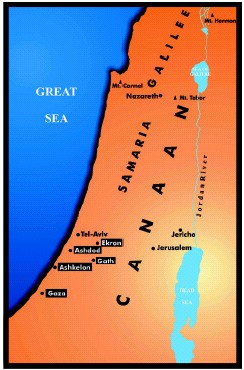
FIGURE 1 Canaan in ancient times. Palestinians (Philistines)
city-states by 1100-1000 BC (black squares) Jericho was an ancient Canaanite
city-state. Jerusalem, Nazareth and modern Tel-Aviv are also indicated
[37]. Palestinian citystates may come from the remains of Egyptian garrisons,
left to their own fate according to archaeological records [6]; the Bible
quotes that Palestinians may have come from Crete [7]. Gath may be placed
more to the south because it has not been recognized in modern times. Goliath,
the Palestine, who was killed by King David according to the Bible, come
from this city.
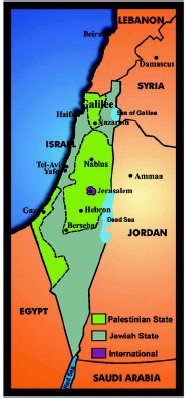
FIGURE 2 United Nations agreement for Palestine partition
in 1947. 890
Palestinians held five important city-states when the fighting with
Israel began (after 1200 BC): Gaza, Ashkelon, Ashod, Ekron and Gath (Figure
1). They won several battles and the time that they were in control of
all Canaan, west to the Jordan river (about 100 years?) has been enough
to name the land as Palestine until after World War II [9]. They probably
had an iron technology higher than Israelites, because they did not let
Israelites to work as blacksmiths when they were ruling over Israelites
[see Bible starting in Samuel 13 and 19; 5,7].
Alexander the Great [9] surrendered Gaza after a long siege about 333
BC. Later, Gaza became an important Christian center [9] and afterwards
an important Islamic center because Palestine was converted to Islam by
Arabic troops and priests by 700 AD [9]. Ancient Canaan (Palestine in Middle
Age and modern times) has sequentially belonged to the Roman-Byzantine
empire, to Egyptian Muslim Mammeluks, to the European crusaders and finally
to the Ottoman-Empire [9] since the XVI century. In 1918 British led mixed
Arab-British troops seized the region. Palestine had 750,000 inhabitants
in 1919 [9] and only 70,000 were Jewish. Immigration rapidly increased
the number of Jews (who had been several times led to Diaspora, expelled,
deported, and massacred by ancient Iranians and Romans, most western European
countries, and finally Hitler [8,9]). There were 400,000 Jews present in
Palestine by 1936 and 600,000 by 1947 when Palestine population amounted
2,000,000 inhabitants. The United Nations plan for Palestine partition
in 1947 is illustrated in Figure 2 [9]. Israel´s self-proclaimed
independent in 1948 and started a war against Muslim Palestinians and other
Muslim neighbouring countries. After several regional wars, Israel has
taken more space and sized Jerusalem, as illustrated in Figure 3. The present
situation (April 2001) is unstable.
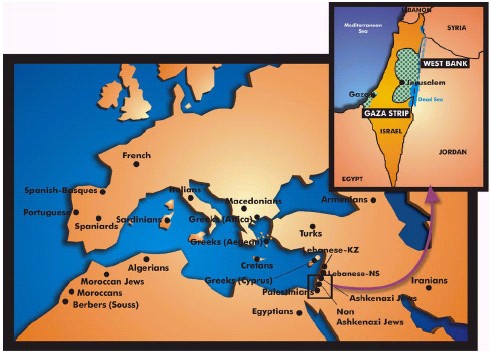
FIGURE 3 Location of the studied populations and map
of the present day Israel-Palestine. The Palestinian Autonomous Government
has a limited rule in the Gaza Strip and West (Jordan) Bank. 891
Palestinians were about 5,000,000 at the beginning of the last decade.
Nowadays, they might reach 7,000,000 [9-11]. Three kind of Palestinians,
according to their status, may be defined as follows: (1) the inhabitants
under an autonomous Palestinian government (about 30%) that controls very
little of the West Jordan Bank (inhabited by more than 1,500,000 Palestinians),
but controls more in the Gaza strip (about 1,000,000 Palestinians), where
Palestinians have to lived mixed with Jewish colonist in their theoretically
own territories; (2) the Diaspora Palestinians (occurred after 1947), who
have refugee status (about 40%, [9,10]), and live either in concentration
camps or are scattered in Jordan (38%), Syria (12%) and Lebanon (13%).
Saudi Arabia and Kuwait gather about 500,000 Palestinians; Egypt, Iraq
and other Mediterranean, European and American countries have more reduced
communities; and (3) the Israel Palestinians, who live within Israel (37%).
Most of Palestinians profess Muslim religion, but there are also Druze
and Christian minorities [10]. They speak the Palestinian-Arab dialect
[10].
The aim of the present study is to examine the genetic relationships
of Palestinians with their neighbours (particularly the Jews) and other
Mediterranean populations in order to: (1) discover the Palestinian origins,
and (2) explain the historic basis of the present day Middle East conflict
between Palestinians and other Muslim countries with Israelite Jews.
MATERIAL AND METHODS
Population samples
Samples from 165 unrelated Palestinians in Gaza (Laboratories and Blood
Bank, El-Shifa Hospital, Gaza) were used for HLA genotyping and phylogenetic
calculations. All were selected in order that their ancestors (eight grandparents)
had a Palestinian origin. This sample may be considered as representative
of the Palestinian population because many people have been forced to live
in the Gaza strip, coming from other parts of Palestine, and the Gaza strip
is now a relatively or totally (when the ongoing conflict aggravates) secluded
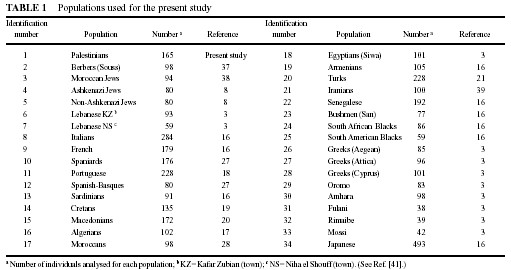
area. All other populations used for comparisons are detailed in Table
1 and Figure 3.
HLA Genotyping, DNA Sequencing and Statistics
Generic HLA class I (A and B) and high resolution HLA class II (DRB1
and DQB1) genotyping was performed using a reverse dot-blot technique with
the Automated Innolipa system (Innogenetics N.V., Zwijndrecht, Belgium).
HLA-A, -B, -DRB1, and -DQB1 allele DNA sequencing was only done when indirect
DNA typing (reverse dot-blot) yielded ambiguous results [12]. Statistical
analysis was performed with Arlequin v1.1 software kindly provided by Excoffier
and Slatkin [13]. In summary, this program calculated HLA-A, -B, -DRB1
and -DQB1 allele frequencies, Hardy-Weinberg equilibrium and the linkage
disequilibrium between two alleles at two different loci. Linkage disequilibrium
(D´; also named LD, see Imanishi et al. [14]) and its level of significance
(p) for 2 x 2 comparisons were determined using the formulae of Mattiuz
and co-workers [15] and the 11th International Histocompatibility Workshop
methodology [14].
In addition, the most frequent complete haplotypes were deduced following
a methodology used in the 11th International Histocompatibility Workshop:
(1) the 2, 3, and 4 HLA loci haplotype frequencies [3,16-21]; (2) the haplotypes
previously described in other populations [3,16]; and (3) haplotypes that
were assigned if they appeared in two or more individuals and the alternative
haplotype was well defined. In order to compare allelic and haplotype HLA
frequencies with other populations, the reference tables used were those
of the 11th and 12th International HLA Workshops [3,16]; see also Table
1. Phylogenetic trees (dendrograms) were constructed with the allelic frequencies
by applying the Neighbor-Joining (NJ) method [22] with the genetic distances
between populations (DA, [23]) and using DISPAN software containing the
programs GNKDST and TREEVIEW [24,25]. A three-dimensional correspondence
analysis and its bidimensional representation was carried out using the
VISTA v5.02 computer program ([26]; http:/
forrest.psych.unc.edu). Correspondence analysis comprises a geometric
technique that may be used for displaying a global view of the relationships
among populations according to HLA (or other) allele frequencies. This
methodology is based on the allelic frequency variance among populations
(similar to the classical principal components methodology) and on the
display of a statistical projection of the differences.
RESULTS
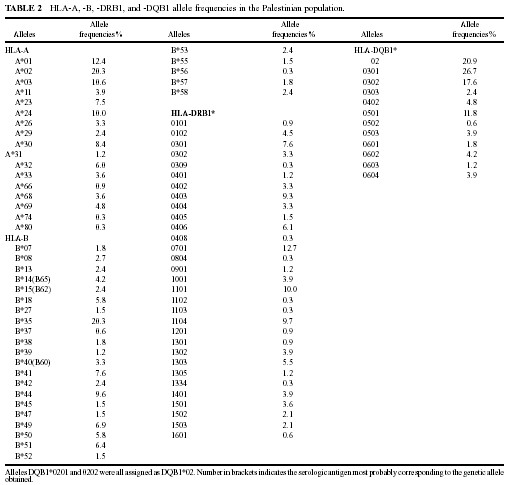
Characteristic HLA Allele Frequencies of the Palestinian Population
Compared With Other Mediterraneans The expected and observed gene allelic
frequencies for HLA-A, -B, -DRB1 and -DQB1 loci do not significantly differ
and the population sample is in Hardy-Weinberg equilibrium. Table 2 illustrates
the HLA allele frequencies found in the Palestinian population. Seventeen
HLA-A and 26 different HLA-B alleles were observed in the Palestinian population.
Seven HLA-A and seven HLA-B alleles had frequencies higher than 5% (A*01,
A*02, A*03, A*23, A*24, A*30, A*32, B*18, B*35, B*41, B*44 B*49, B*50,
and B*51) and these are characteristic of Mediterranean populations, particularly
from eastern Mediterranean regions [17,19- 21,27,28]. With regard to the
HLA class II alleles, 31 different DRB1 alleles were found and only seven
had frequencies higher than 5% (DRB1*0301, *0403, *0406, *0701, *1101,
*1104, *1303) being also characteristic of eastern Mediterranean populations.
In particular, DRB1*0403 is present in high frequency in Lebanese [29],
North African and Jewish populations [8,17]; DRB1*0406 is also present
in North Africans populations [17,28]. Besides, DRB1*0302 (allele frequency
3.3%), characteristic of African Black populations, is also present in
North African Caucasians, Jews and Lebanese [8,17,28,29]. DQ allele frequencies
reflect the DRB1 locus allele distribution due to the strong linkage disequilibrium
between these two loci. Three types of analyses were carried out to compare
Palestinian HLA frequencies with other Mediterranean population frequencies:
(1) with DRB1-DQB1 data, which is probably a more informative and discriminating
methodology; (2) with DRB1 data; and (3) with generic (low resolution)
DR-DQ data. These three types of analysis were performed because some of
the populations used for comparison lacked HLA-A and -B data (Berbers [from
Souss, Agadir area], Jews [Ashkenazi], Jews [Morocco], Jews [non-Ashkenazi],
Lebanese [NS and KZ], see Table 1), or high resolution HLA-DQ data (Greeks
[Attica], Greeks [Cyprus], Greeks [Attica- Aegean], see Table 1), or only
generic HLA-DR and DQ data were available (Portuguese, Turks, Iranians,
Armenians and Egyptians, see Table 1). These partially HLA-typed populations
should have been ignored, but they could be analyzed conjointly taking
into account only either DRB1 or generic DR and DQ frequencies (Table 3,
Figures 1, 2, 3, 4 and 5).
Analyses using DRB1 and DQB1 conjointly were made but are not illustrated
because only a few populations could be used and the results are concordant
with the DRB1 analysis. Finally, it should be pointed out that class I
generic typing tends to homogenize the comparisons based on DRB1 high resolution
typing [28]; one class I allele obtained by generic DNA typing may contain
several class I alleles, whereas this is not the case for most DRB1 alleles
at present.
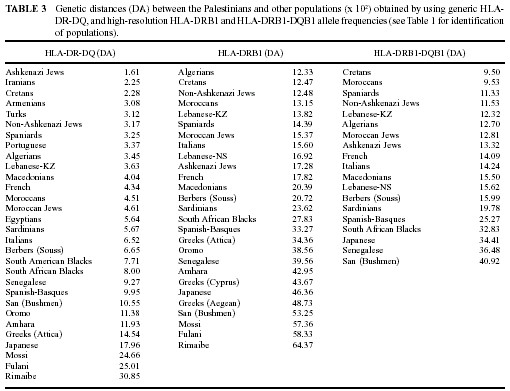
Figure 4 depicts an HLA class II high-resolution (DRB1) NJ tree. Populations
are grouped into three main branches: the first one groups both eastern
(including Palestinians, Macedonians, Cretans, Jews, Lebanese) and western
Mediterranean populations (Europeans and North Africans; Sardinians are
included in the first group). The second branch is formed by African Negroid
populations and Japanese (Mediterranean outgroups) and the third one includes
Greek and Ethiopian/sub-Saharan populations. This distribution is also
confirmed in the correspondence analysis (Figure 5): the three groups are
clearly delimited and an smooth West to East Mediterranean gradient is
shown. The Palestinian population reveals the closest genetic distance
with Jews (generic DR-DQ typing), Cretans (using DRB1-DQB1) or Algerians
(using DRB1) (see Table 3) and no discontinuity is observed between eastern
and western Mediterranean populations reflecting the genetic similarity
among all these populations. It is evidenced that Palestinians/Greeks distance
is high and confirms the different genetic background of the Greeks, who
have received a substantial sub-Saharan gene flow [3,20]. These results
are confirmed using DR and DQ generic typings (see Table 3 and Figure 6)
which were used in 893 order to include other Mediterranean populations
(Iranians, Armenians, Egyptians and Turks, see Table 1). The close relatedness
of Palestinians (Table 3 first column, Figure 6) to Iranians, Armenians,
Egyptians and Anatolians (Turks [21]) further support an autochthonous
Canaanite/Middle East origin for both Palestinians and Jews. A DR-DQ neighbour-joining
tree (not shown) maintains the west and east Mediterranean groups and also
the group formed by Greeks and sub-Saharan populations. Turks (old Anatolians),
Armenians, Jews and Lebanese are illustrated specifically to cluster with
Palestinians. On the other hand, genetic distances obtained by using DR-DQ
generic typing allele frequencies (Table 3) illustate that Ashkenazi Jews,
Iranians, Cretans, Armenians, Turks and non-Ashkenazi Jews are the populations
closest to the Palestinians, followed by the other Mediterraneans populations.
Other analyses and genetic distances confirm these results (Table 3, Figures
4 and 5).
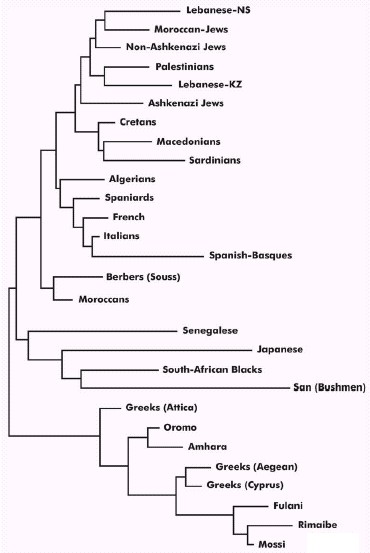
FIGURE 4 Neighbor-Joining dendrogram demonstrating
relatedness between Palestinians and other populations. Genetic distances
between populations (DA) were calculated by using HLA-DRB1 (high resolution).
Data from other populations were from references detailed in Table 1. Bootstrap
values from 1000 replicates are illustrated. 896
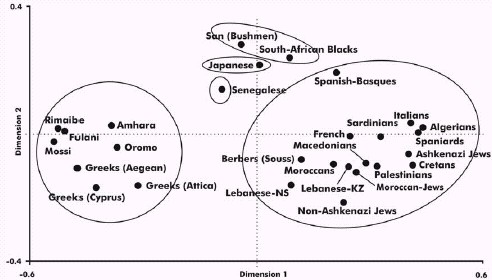
FIGURE 5 Correspondence analysis depicting a global
view of the relationship between Mediterraneans and Palestinians according
to HLA allele frequencies in three dimensions (bidimensional representation).
HLA-DRB1 allele frequencies data. 897
HLA-A, -B, -DRB1, and -DQB1 Linkage Disequilibria in Palestinians Extended
HLA haplotypes were defined in Palestinians and compared with those previously
reported in other populations (Table 4).

TABLE 4 Most frequent HLA-A, -B, -DRB1, and -DQB1 extended
haplotypes in the Palestinian population and their possible origin.
HLA-A-B and DRB1*-DQB1* two-loci linkage disequilibrium data (not shown)
show that the most frequent combinations are characteristic of Mediterranean
(western and eastern) populations (A*1-B*35, haplotype Frequency (HF):
5.1; B*35-DRB1*1104, HF: 3.2; B*35-DRB1*0403, HF: 3.2; A*2-B*41, HF: 3.1;
A*2-B*51, HF: 2.9; B*18-DRB1*1104, HF: 2.8; B*49-DRB1*0403, HF: 2.7; A*24-B*35,
HF: 2.3; A*23-B*44, HF: 2.3; A*24-B*18, HF: 2.1; B*14- DRB1*0102, HF: 2.0;
B*35-DRB1*1101, HF: 2.0; A*33-B*14, HF: 1.7; B*49-DRB1*1001, HF: 1.6; B*35-DRB1*1001,
HF: 1.5; B*50-DRB1*0701, HF:1.5; B*8-DRB1*0301, HF: 1.5). The combination
A*69-B*49 (HF: 2.7) has not been found in any of the populations tested
and it is included in an extended haplotype (A*69-B*49-DRB1*0403-DQB1*0302,
see below) not previously described.
The HLA-A-B-DR-DQ extended haplotypes found in the Palestinian population
(Table 4) reflect common characteristics with the other older Mediterranean
population mainly from eastern Mediterraneans and North Africans (see footnote
to Table 4), like Jews [8]. These haplotype results are concordant with
those obtained by the allele frequency analyses (genetic distances, NJ
trees and correspondence analyses, see above).
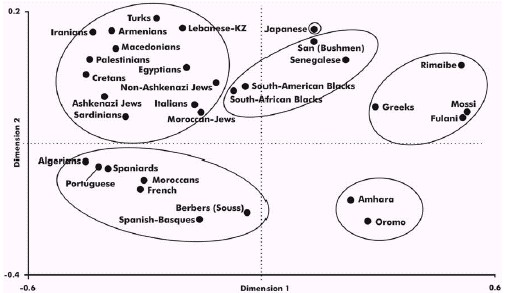
FIGURE 6 Correspondence analysis depicting a global
view of the relationship among Palestinians, West Mediterraneans, East
Mediterraneans, Greeks and sub- Saharian populations and Blacks according
to HLA allele frequencies in three dimensions (bidimensional representation)
. HLA-DR and DQ (low resolution) allele frequencies data. 898
DISCUSSION
Palestinians and Jews The genetic identity of Ashkenazi and non-Ashkenazi
Jews who now lives in Israel has already been reported [8]. Babylonian
and Roman-induced Diaspora, drove Jews to many parts of Europe, Africa
and Asia, which occurred in 587 BC and 70 AD, respectively. Jews started
to come back to Palestine during the 19th and 20th centuries [8]. However,
religion and close communities have kept Jews relatively isolated from
the inhabitants of the countries that hosted them during this long period
of time. Jews wrote the Bible, a religious and historical book that is
a continuous source of historical Middle East facts, but that only tells
the Jewish view [6]. It is now necessary to rely on other sources, such
as archaeology, linguistics, etc, to establish a more objective history
of Middle East and particularly ancient Canaan [6,9].
Palestinians appeared in the Bible as coming from Crete or its empire
[7]. The present day concept based in archaeology is that most original
Palestinians were already in Canaan and some tribes were agglutinated by
Egyptian garrisons, left to their own fate in Canaan [6]; but the input
of one elite coming from Crete may not be discarded. Also, the bulk of
Jewish people probably came from ancient autochthonous Canaanites [6];
this is compatible with an input of foreign leaders and their groups (Abraham,
Moses) as described in the Bible [7].
Both Jews and Palestinians share a very similar HLA genetic pool (Table
3, Figures 4, 5 and 6) that support a common ancient Canaanite origin.
Therefore, the origin of the long-lasting Jewish-Palestinian hostility
is the fight for land in ancient times. Religious and cultural have enhanced
the conflict in the last centuries, together with the massive European,
American, Asian and African Jews settlements in the area, which has also
caused a massive displacement of Palestinians and wars. A difficult problem
has now been created between two communities that are close genetic relatives.
Regarding Palestinian population identity, it is clear that they spoke
a language different to Arab or Jewish in ancient times and only a few
words have been preserved. Palestinians named their leaders or princes
as seren (Basque, Zar = old man, en = the most important) [7]. The study
of this and other words suggests that they spoke a Dene-Caucasian language
like other Mediterranean populations [30,31]. The typical Philistine crest-hut
already appeared in the Cretan Phaistos Disk (1600 BC) and in the Ramses
III-Medinet Habu temple, Egypt (1200 BC, [5]).
The Eurocentric confusion Arab = Muslim has also lowered the Palestinian
identity by identifying the country were Mohammed was born (Saudi Arabia)
with the Muslim religion; it also has artificially divided peoples both
coming from ancient Canaanites (Jews and Palestinians).
Palestinians and Other Middle East and European People Palestinians
are close to Egyptians, Lebanese, Iranians, Cretans, Macedonians and Sardinians,
and also to Algerians, Spaniards, French, Italians and Basques (Table 3,
Figures 4, 5, and 6). DRB1 genetic distances (Table 1) are probably the
most reliable ones due to the higher polymorphism detected in this locus.
The western and eastern Mediteranean populations are intermingled in this
case; it supports the long-standing prehistoric and historic circum-Mediteranean
gene flow [32]. Jews, Cretans, Egyptians, Iranians, Turks and Armenians
are probably the closest relatives to Palestinians and this favors the
hypothesis that most of the HLA Palestinian genetic background comes from
the Middle East (ancient Canaan, [6]), ancient stock, i.e.: ancient Canaanites.
Canaan had received gene and cultural flow from Mesopotamia, Anatolia
and Egypt [6].
Palestinians, Cretans and Greeks The Biblical origin for Palestinians
(Crete) cannot be disregarded [7] because an elite group could have joined
to Canaanite proto-Palestinian tribes and made themselves noticeable; this
is supported by the ancient Palestinians high war technology and the many
confrontations with the Jews after 1500 BC [6,7]. It is very unlikely that
a massive immigration of Palestinians came from Crete [6]. Egyptian garrisons
in Canaan abandoned to their fate by the Egyptian Kingdom weakness may
have catalyzed the union of some Canaanite tribes to become the historical
Palestinians, according to Amelie Kuhrt [6].
By 1500-1200 BC the Greek presence was very scarce in Canaan, according
to archaeologic records [6]. In fact, the Mycaenian Greeks attacked Crete
by 1450 BC after rendering tributes to Cretans by a relatively long period.
The Cretan Aegean Sea empire was destroyed and continued by the Mycaenians.
Greeks are found to have a substantial HLA gene flow from sub-Saharan Ethiopian
and Black people [3,20]. This is why Greeks are Mediterranean outliers
in all kind of analyses [19-21,28]. This African genetic and cultural input
was documented by Herodotus [33] who states that the daughters of Danaus
(who were black) came from Egypt in great numbers to settle in Greece.
Also, ancient Greeks believed that their religion and culture came from
Egypt [33]. An explanation of the Egypt-to-Greece migration may be that
a densely populated Sahara (before 5000 BC) may have contained an admixture
of Negroid and Caucasoid populations, and some of the Negroid populations
may have migrated by chance or unknown causes towards present day Greece
[19,34-36].
This could have occurred when hyperarid Saharan condition become established
and large-scale migration occurred in all directions out from the desert.
In this case, the most ancient Greek Pelasgian substratum would come from
a Negroid stock. A more likely explanation is that at an undetermined time
during Egyptian pharaonic times a Black dynasty with their followers were
expelled and went towards Greece where they settled [20, 30].
Once an African input to the ancient Greek genetic pool is established,
it remains to be determined what the cultural importance of this input
is for constructing the classical Hellenistic culture. The reason why a
sub- Saharan admixture is not seen in Crete is unclear but may be related
to the influential and strong Minoan empire, which hindered foreigners
establishment if the African invasion occurred in Minoan times [19, 20].
ACKNOWLEDGMENTS
We are grateful to Alberto Garcia for his help with art design work
on the computer. We also thank M. Nei for providing the GNKDST and TREEVIEW
programs found on the DISPAN software. This work was supported in part
by grants from the Spanish Ministry of Education (PM95-57 and PM96-21)
and the Madrid Regional Government (06/70/97 and 8.3/14/98).
REFERENCES
==1. Dausset J, Colombani J: Histocompatibility Testing 1972. Copenhagen,
Munksgaard, 1972.
==2. Imanishi T, Wakisaka A, Gojorobi T: Genetic relationships among
various human populations indicated by MHC polymorphisms. In Tsuji K, Aizawa
M, Sasazuki T (eds): HLA 1991. Vol 1. Oxford, Oxford University Press,
627-32, 1992.
==3. Clayton J, Lonjou C: Allele and Haplotype frequencies for HLA
loci in various ethnic groups. In Charron D (ed): Genetic diversity of
HLA, Functional and Medical Implications. Vol 1. Paris, EDK, 665-820, 1997.
==4. Encyclopaedia Britannica (1999) Sutton, Surney: Encyclopaedia
Britannica International Ltd.
==5. Kienitz FK: Volker imm schatten. Die gegenspieter der Griechen
und Römer. Munchen, Becksche Verlagsbuchhandlung, 1981.
==6. Kuhrt A: The ancient Near East (3000-330 BC). Vol II. Barcelona,
Editorial Critica, 2001.
==7. Biblia de Jerusalén. Desclee de Brouwer (ed): Bilbao, Spain,
1998.
==8. Martínez-Laso J, Gazit E, Gómez-Casado E, Morales
P, Martinez-Quiles N, Alvarez M, Martin-Villa JM, Fernandez V, Arnaiz-Villena
A: HLA DR and DQ polymorphism in Ashkenazi and non-Ashkenazi Jews: comparison
with other Mediterraneans. Tissue Antigens 47: 63, 1996.
==9. Sellier J, Sellier A: Atlas des Peuples dOrient. Paris, Editions
La Dècouverte, 1993.
==10. Gonem A: The encyclopaedia of the peoples of the world. Jerusalem,
Publishing House Ltd, 143-5, 1996.
==11. El Estado del Mundo, 2000. Anuario geopolítico. Madrid,
Akal Editions, 2000.
==12. Arnaiz-Villena A, Timón M, Corell A, Pérez-Aciego
P, Martín-Villa JM, Regueiro JR: Primary immunodeficiency caused
by mutations in the gene encoding the CD3-gg subunit of the T-lymphocyte
receptor. N Eng J Med 327:529, 1992.
==13. Excoffier L, Slatkin M: Maximum-likelihood estimation of molecular
haplotype frequencies in a diploid population. Mol Biol Evol 12:921, 1995.
==14. Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojorobi T: Estimation
of allele and haplotype frequencies for HLA and complement loci. In Tsuji
K, Aizawa M, Sasazuki T (eds): HLA 1991. Vol I. Oxford, Oxford University
Press, 76-9, 1992.
==15. Mattiuz PL, Ihde D, Piazza A, Ceppelini R, Wodmer WF. Histocompatibility
Testing 1970. Copenhagen, Munksgaard, 193-206, 1970
==16. Imanishi T, Akaza T, Kimura A, Tokunaga K, Gojobori T: Allele
and haplotype frequencies for HLA and complement loci in various ethnic
groups. In Tsuji K, Aizawa M, Sasazuki T (eds): HLA 1991. Vol 1. Oxford:
Oxford University Press, 1065-220, 1992.
==17. Arnaiz-Villena A, Benmamar D, Álvarez M, Díaz-
Campos N, Varela P, Gomez-Casado E, Martinez-Laso J: HLA allele and haplotype
frequencies in Algerians. Relatedness to Spaniards and Basques. Hum Immunol
43:259, 1995.
==18. Arnaiz-Villena A, Martínez-Laso J, Gómez-Casado
E, Díaz-Campos N, Santos P, Martinho A, Breda-Coimbra H: Relatedness
among Basques, Portuguese, Spaniards, and Algerian studied by HLA allelic
frequencies and haplotypes. Immunogenetics 47:37, 1997.
==19. Arnaiz-Villena A, Iliakis P, González-Hevilla M, Longás
J, Gomez-Casado E, Sfyridaki K, Trapaga J, Silvera-Redondo C, Matsouka
C, Martínez-Laso J: The origin of Cretan population as determined
by characterization of HLA alleles. Tissue Antigens 53:213, 1999.
==20. Arnaiz-Villena A, Dimitroski K, Pacho A, Moscoso J, Gomez-Casado
E, Silvera C, Varela P, Martinez-Laso J: HLA genes in Macedonians and the
sub-Saharan origin of the Greeks. Tissue Antigens 57:118, 2001.
==21. Arnaiz-Villena A, Carin M, Bendikuze N, Gomez- Casado E, Moscoso
J, Silvera C, Pacho A, Allende L, Guillén J, Martinez-Laso J: HLA
alleles and haplotypes in the Turkish population: relatedness to Kurds,
Armenians and other Mediterraneans. Tissue Antigens 57:308, 2001.
==22. Saitou N, Nei M: The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Mol Biol Evol 4:406, 1987.
==23. Nei M: Genetic distances between populations. Am Nat: 106:283,
1972.
==24. Nei M: Analysis of gene diversity in subdivided populations.
Proc Natl Acad Sci USA 70:3321, 1973.
==25. Nei M, Tajima F, Tateno Y: Accuracy of estimated phylogenetic
trees from molecular data II. Gene frequency data. J Mol Evol 19:153, 1983.
==26. Young FW, Bann CM: A Visual Statistics system. In Stine RA &
Fox J (eds): Statistical Computing Environments for Social Researchers.
New York, Sage publications, 207-36, 1996.
==27. Martínez-Laso J, De Juan D, Martínez-Quiles N,
Gómez-Casado E, Cuadrado E, Arnaiz-Villena A: The contribution of
the HLA-A, -B, -C and -DR, -DQ DNA typing to the study of the origins of
Spaniards and Basques. Tissue Antigens: 45:237, 1995.
==28. Gomez-Casado E, del Moral P, Martinez-Laso J, García-
Gómez A, Allende L, Silvera-Redondo C, Longas J, González-Hevilla
M, Kandil M, Zamora J, Arnaiz- Villena A: HLA genes in Arabic-speaking
Moroccans: close relatedness to Berbers and Iberians. Tissue Antigens 55:239,
2000.
==29. Bias W, Gazit E: 12th International Histocompatibility Workshop:
Anthropology SWAS regional report. In Charron D (ed): Genetic diversity
of HLA, Functional and Medical Implications. Vol 1. Paris, EDK, 353-363,
1997.
==30. Arnaiz-Villena A, Martinez-Laso J, Alonso-García J: The
correlation between languages and genes: the usko- Mediterranean peoples.
Human Immunol (This issue)
==31. Arnaiz-Villena A, Alonso-Garcia J: The Usko-Mediterranean languages.
In Arnaiz-Villena A (ed): Prehistoric Iberia: Genetics, Anthropology and
Linguistics. New York, Kluwer Academic-Plenum Publishers, 2000.
==32. Arnaiz-Villena A, Martinez-Laso J, Alonso-Garcia A. Iberia: Population
genetics, Anthropology, and linguistics. Hum Biol 71:725, 1999.
==33. Herodotus: History. Madrid, Ed. Gredos, 1989.
==34. Arnaiz-Villena A, Alonso-García J: El origen de los Vascos
y otros pueblos Mediterraneos. Madrid, Editorial Complutense SA, 1998.
==35. Arnaiz-Villena A, Alonso-García J: Minoicos, Cretenses
y Vascos. Un estudio genético y lingüístico. Madrid,
Editorial Complutense SA, 1999.
==36. Arnaiz-Villena A, Alonso-García J: Egipcios, Bereberes,
Guanches y Vascos. Madrid, Editorial Complutense SA, 2000.
==37. Grollemberg LH: The Penguin shorter Atlas of the Bible. New York,
Penguin Books Ltd, 1978.
==38 Izaabel H, Garchon HJ, Caillat-Zucman S, Beaurain G, Akhayat O,
Bach JF, Sanchez-Mazas A: HLA class II DNA polymorphism in a Moroccan population
from the Souss, Agadir area. Tissue Antigens 51: 106, 1998.
==39. Roitberg-Tambur A, Witt CS, Friedmann A, Safirman C, Sherman
L, Battat S, Nelken D, Brautbar C: Comparative analysis of HLA polymorphism
at the serologic and molecular level in Moroccan and Ashkenazi Jews. Tissue
Antigens 46: 104, 1995.
==40. Mehra NK, Rajalingam R, Kanga U, McEnemy L, Cullen C, Agarwal
S, Middleton D, Pollack MS, Amirzargar A, Singal DP: Genetic diversity
of HLA in the populations of India, Sri Lanka and Iran. In Charron D (ed):
Genetic diversity of HLA, Funcional and Medical Implications. Vol 1. Paris,
EDK, 314-20, 1997. 900
click here to download PDF verson
of this article
revised: February 2003
revised January 2004mailto: israels-ekklesia@tellme1st.net
goto top.....